Embryogenesis
Following fertilization of the egg cell by a sperm cell, the zygote develops inside the seed. The sequence of development proceeds through histodifferentiation, cell expansion and maturation drying.
The stage of histodifferentiation (referred to as embryogenesis) and is characterized by differentiation of the endosperm and embryo.
This process is similar but unique for monocots, dicots and gymnosperms.
Monocot embryogenesis - corn
Proembryo stage
Monocots have a more complex embryo structure in the mature seed compared to dicots, but early embryo development is similar to dicots.
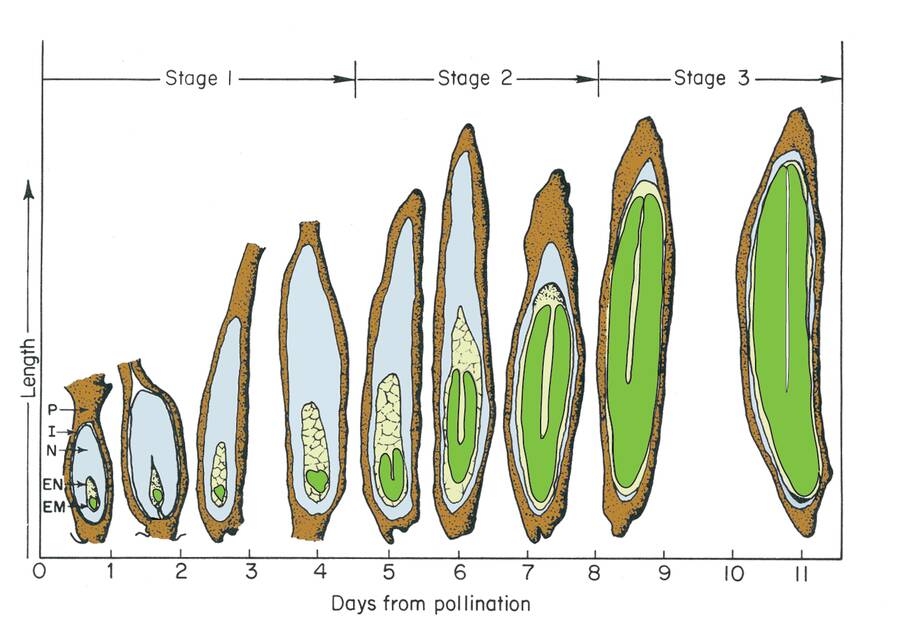
The stages of embryogenesis in monocots include the proembryo, globular, scutellar and coleoptilar stages.
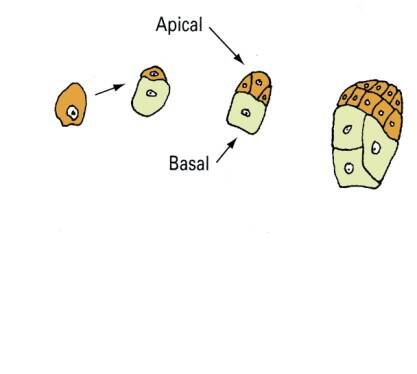
Following fertilization, the first cell division is asymmetrical and leads to an apical and basal cell in corn (Zea mays). The apical cell divides more rapidly than the basal cell and will eventually be the embryo.
Globular stage
The proembryo in the globular stage is similar to dicots, except that the suspensor is not a single or double row of cells and is less differentiated.
In the late globular stage, the outer epidermal layer is evident and a group of cells on one side of the proembryo divides more rapidly.
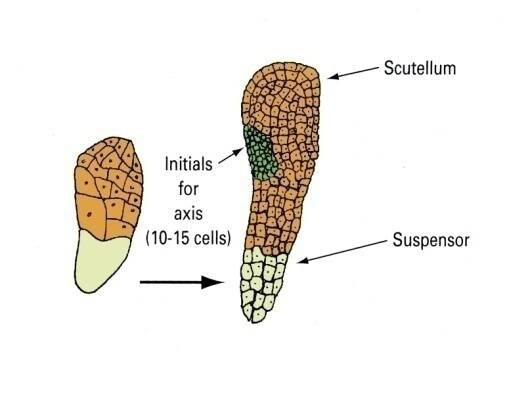
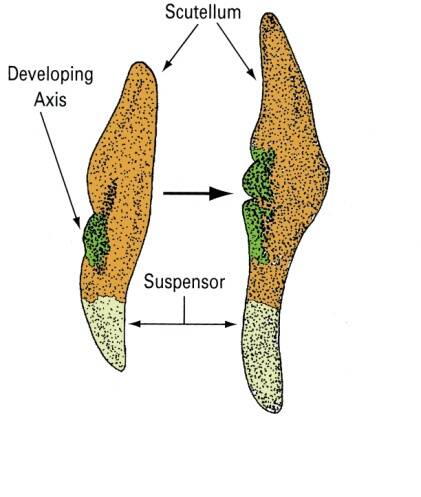
These will give rise to the embryo axis.
Scutellar stage
The remnant of the cotyledon can be seen in the scutellar stage of development.
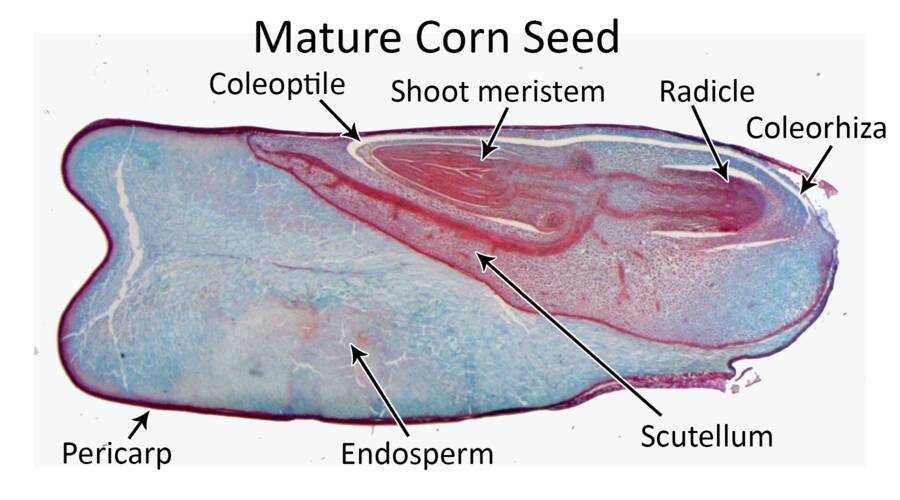
Monocots have reduced the pair of cotyledons represented in dicot embryos to a single modified cotyledon termed the scutellum.
The scutellum acts as conductive tissue between the endosperm and embryo axis.
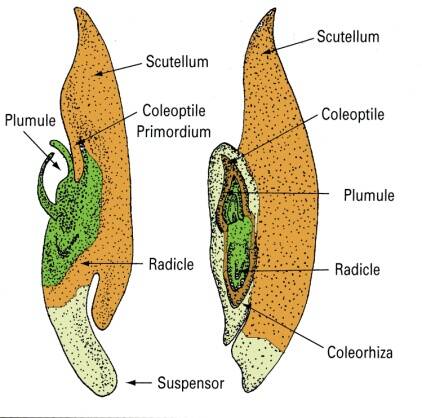
Coleoptilar stage
The embryo axis differentiates into the plumule (shoot) and radicle.
In monocots, the embryo axis also has a specialized tissue surrounding the shoot and root tissue to aid in emergence during germination.
These are the coleoptile and coleorhiza, respectively.
Diocot embryogenesis – Shepherd's purse
Proembryo stage
Shepherd's purse (Capsella bursa- pastoris) serves as a good model for dicot embryogenesis.
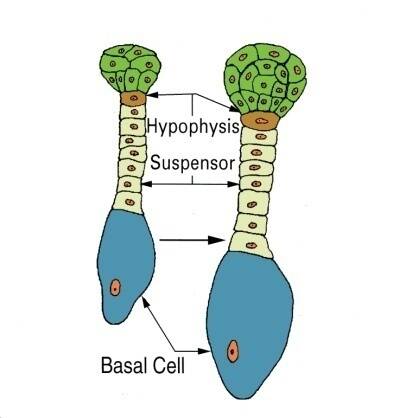
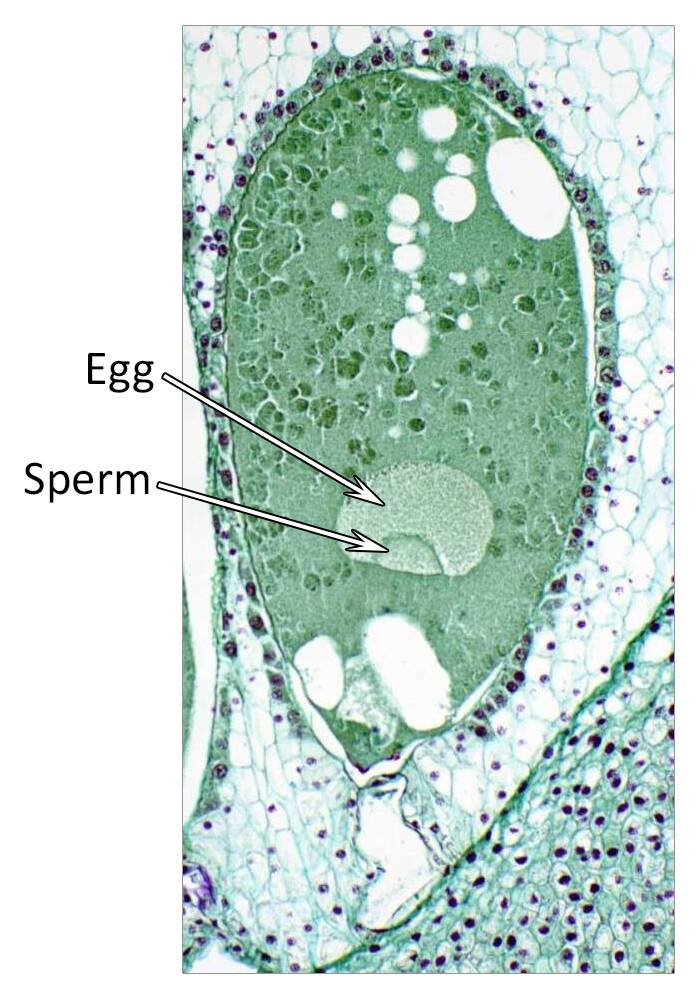
Embryogenesis goes through proembryo, globular, heart, torpedo, and cotyledon stages. Following fertilization of the egg and sperm nuclei, a proembryo is formed by a transverse cell division to form an apical and basal cell.
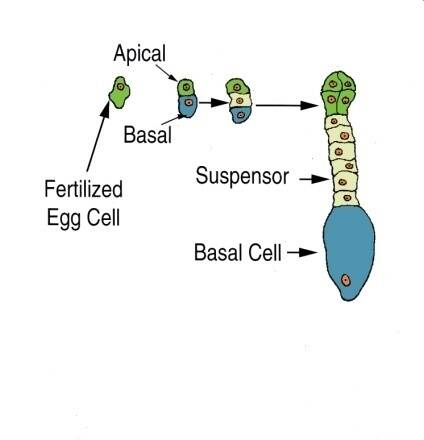
The basal cell forms the suspensor, while the apical cell forms the embryo. The suspensor in dicots is usually a column of single or multiple cells.
The suspensor functions to push the proembryo into the embryo sac cavity and to absorb and transmit nutrients to the proembryo.
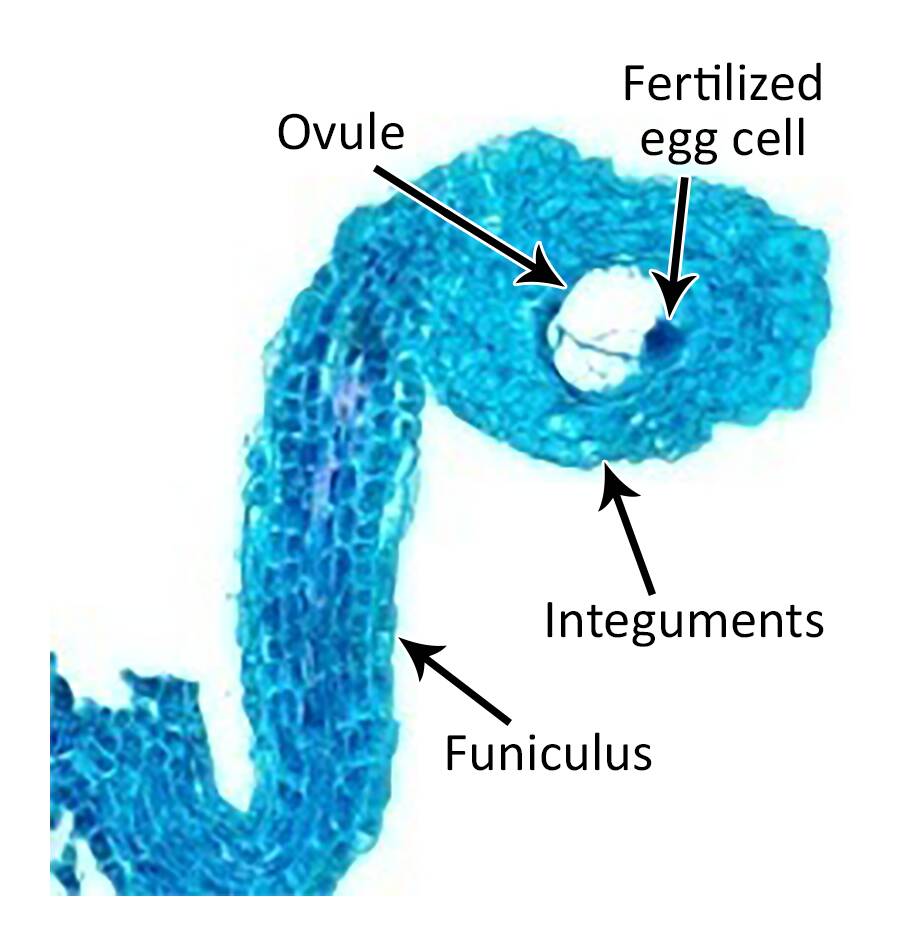
Globular stage
In Shepherd's purse, basal cell derivatives in the globular embryo form the hypophysis that goes on to develop into the radicle. Tissue differentiation becomes evident in the 16-celled globular embryo.
In Shepherd's purse, basal cell derivatives in the globular embryo form the hypophysis that goes on to develop into the radicle.
Tissue differentiation becomes evident in the 16-celled globular embryo.
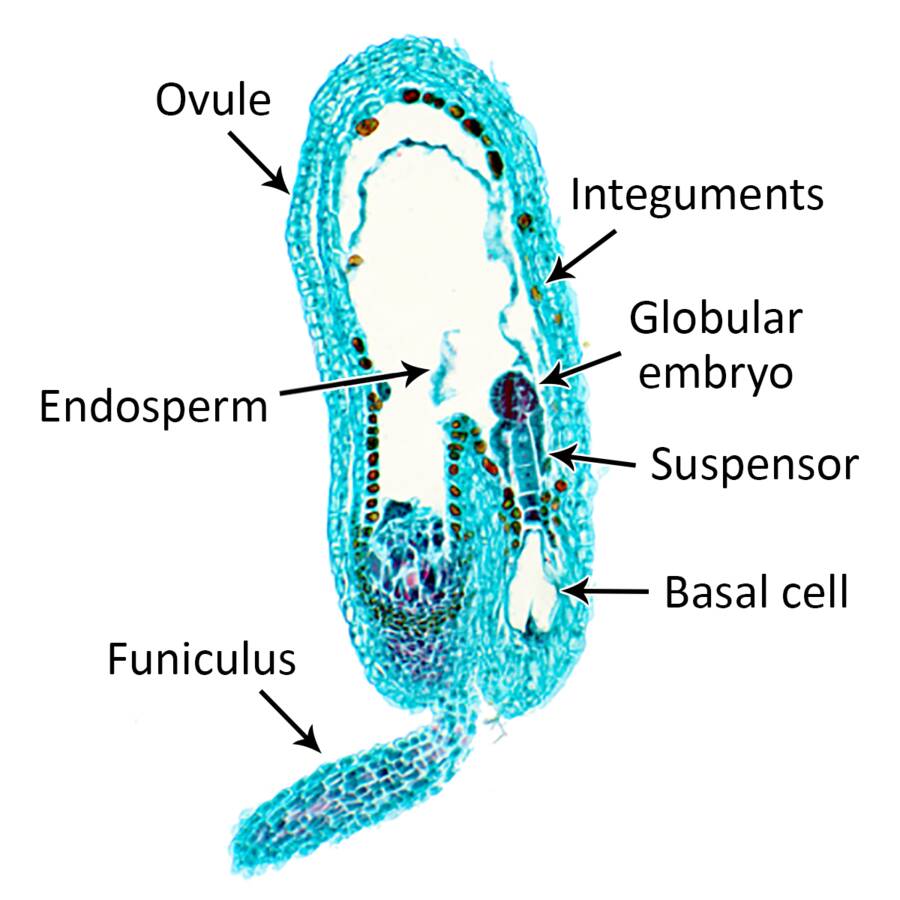
Cotyledon stage
Cotyledon primordium are evident in the heart-shaped stage of embryogenesis.
These primordia elongate to give a typical torpedo stage embryo.
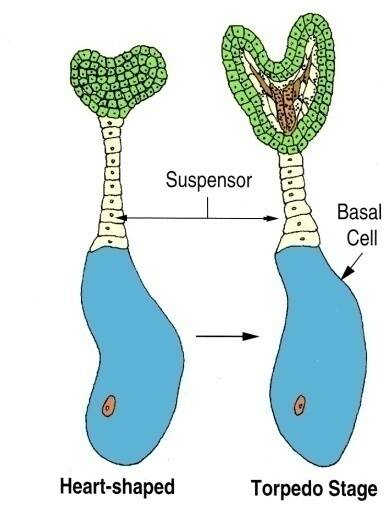
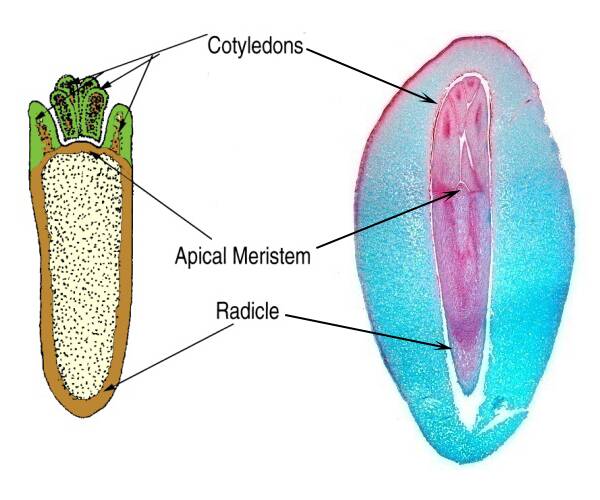
In the heart and torpedo stages, the embryo has organized to form an apical meristem, radicle, cotyledons and hypocotyl.
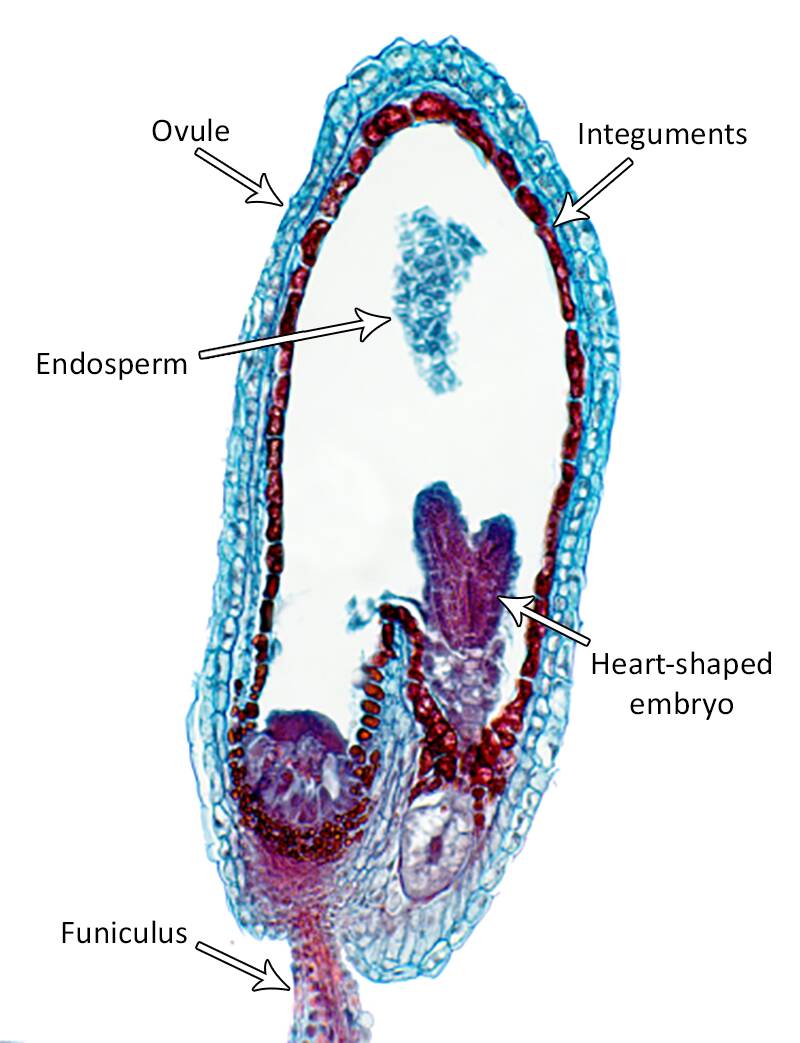
Mature stage
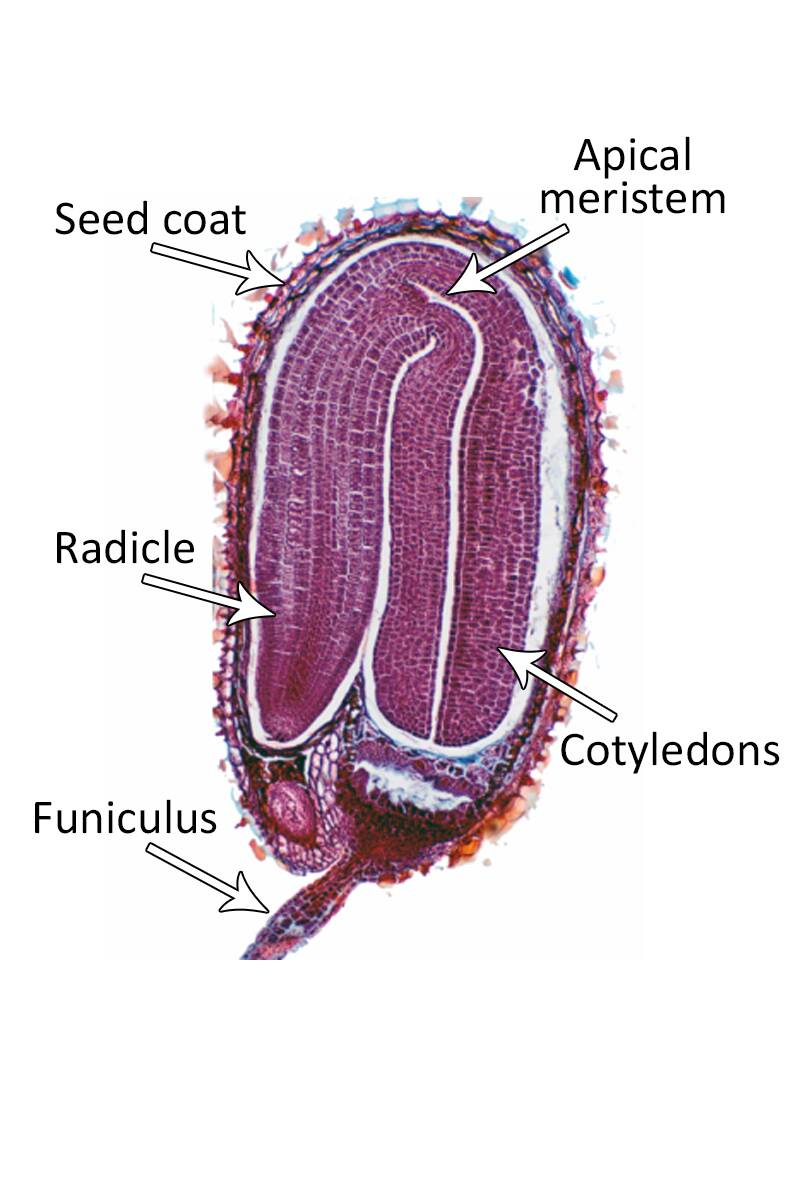
In this stage, the embryo is fully formed and separate cotyledons can be easily seen as well as a distinct shoot and root meristem.
At this time the suspensor and the basal cell begin to disappears.
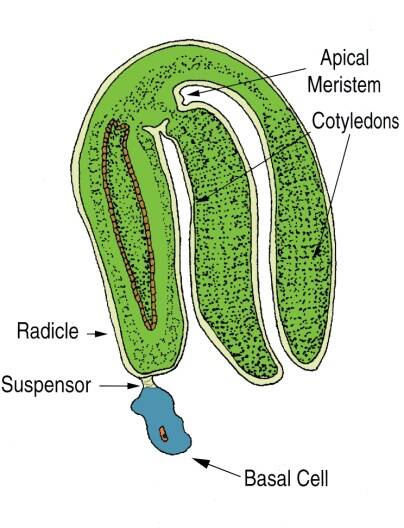
In this stage, the embryo is fully formed and separate cotyledons can be easily seen as well as a distinct shoot and root meristem.
At this time the suspensor and the basal cell begin to disappears.
Gymnosperm embryogenesis – Pine
Compared to the more evolutionarily advanced angiosperms, embryo formation in gymnosperms differs in several important ways.
Most conspicuous is that seeds of gymnosperms are not contained within a carpel (fruit). The term gymnosperm means "naked seeded".
There is also no true triploid endosperm in gymnosperms. Rather, the developing embryo is nourished by haploid female gametophyte tissue.
Free nuclear stage
After fertilization, several embryos begin development within a single gymnosperm seed but rarely does more than one of these embryos mature.
In pine (Pinus sp.), the fertilized egg cell divides to form a free nuclear stage without cell walls between nuclei.
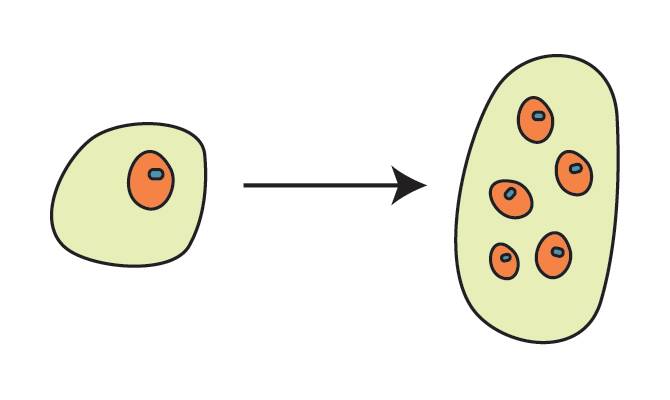
Suspensor tier stage
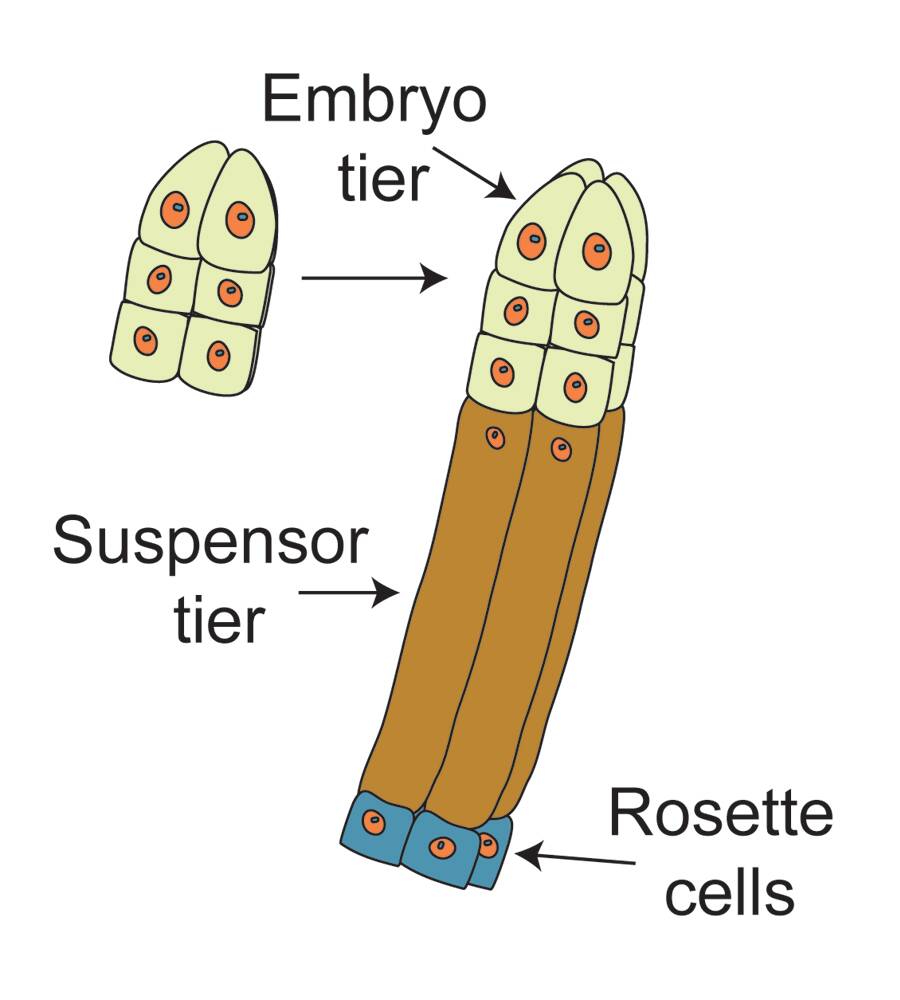
Following cell wall formation, cells organize to form an embryo tier of cells and a suspensor tier.
The suspensor differentiates into a set of primary suspensor cells (rosette cells) and embryonal suspensor tubes.
Proembryo stage
The suspensor cells elongate and there are several cleavage events to give multiple embryos (polyembryos) inside a single seed.
Usually, only one of these embryos continue to develop.
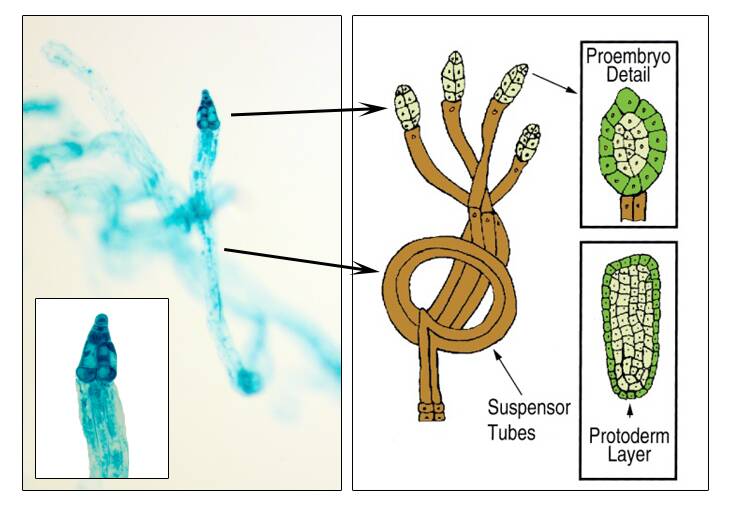
Cotyledon stage
The proembryo continues to differentiate an epidermal layer prior to multiple cotyledon primordia becoming evident in the cotyledon stage.
Also, the shoot and root meristem has been developed.